Regulation of ETV4 expression by mechanical cues
To identify transcriptional regulators that link cell crowding to transcriptional responses in hESCs, we manipulated local cell crowding by limiting the cell adhesive area and performed RNA sequencing (RNA-seq) (Fig. 1a, Extended Data Fig. 1b, and Supplementary Table 1). Cell crowding was verified by measuring the average area of cells covering the substrate (hereafter referred to as cell area; Extended Data Fig. 1a). Gene ontology (GO) analysis with total differentially expressed genes (DEGs) (|log2(fold change)| > 0.5, adjP < 0.05) revealed the top 10 GO terms related to embryonic development and differentiation (Extended Data Fig. 1c)24, supporting the relevance of cell crowding in developmental processes. Contractile actomyosin bundles represent a reliable indicator of cellular mechanical stress levels. Gene set enrichment analysis (GSEA) revealed a significant reduction in genes related to actomyosin in crowded hESCs (Extended Data Fig. 1d)25, confirming decreased mechanical stress. These results were validated by immunostaining for phosphorylated myosin light chain (pMLC) that marks contractile actomyosin bundles (Extended Data Fig. 1e).
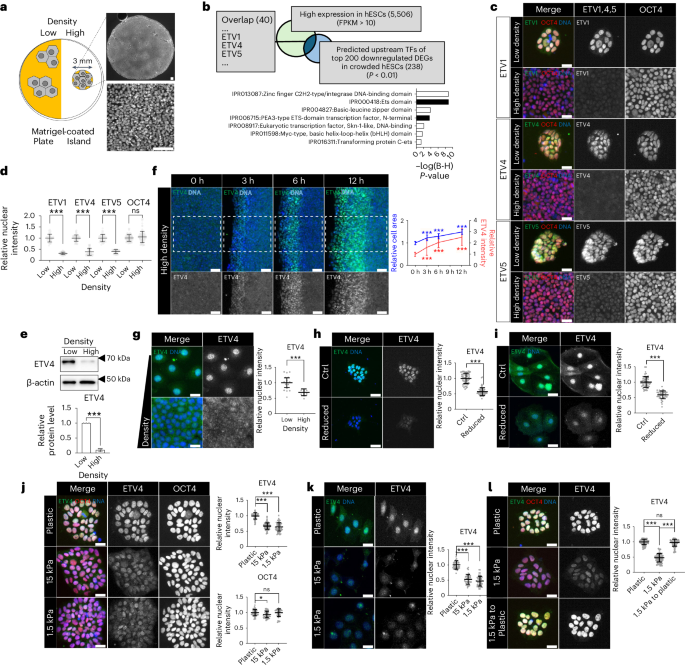
a, Schematic representation of high-density culture. Cell adhesive areas were reduced by matrigel-coated islands. b, Significant terms from InterPro protein domain analysis of 40 predicted upstream regulators of top 200 downregulated DEGs in crowded hESCs with high expression (>10 FPKM). A full list of the 40 proteins can be found in Supplementary Table 2. B-H, Benjamini-Hochberg. c,d, Representative images (c) and quantifications (d) of immunofluorescence assay for ETV1,4,5 and OCT4 in different densities of H9 hESCs. n = 40 cells for ETV1, n = 55 cells for ETV4, n = 45 cells for ETV5, n = 77 cells for OCT4. e, Western blots of ETV4 in different densities of H9 hESCs. n = 3 independent experiments. f, Immunofluorescence assay for ETV4 in scratched H9 hESCs. n = 40 cells for 0 h, n = 50 cells for 3, 6 and 12 h. Cell area was measured by dividing the total surface area by the number of cells. n = 9 regions for relative cell area. g, Immunofluorescence assay for ETV4 in MCF-7 cells. n = 30 cells for low density and n = 41 cells for high density. h, Immunofluorescence assay for ETV4 in H9 colonies on the cell stretching system. n = 60 cells. i, Immunofluorescence assay for ETV4 in MCF-7 cells on the cell stretching system. n = 59 cells for control and n = 46 reduced cells. j, Immunofluorescence assay for ETV4 and OCT4 in H9 hESCs on PDMS layers with different stiffnesses. ETV4: n = 80 cells for plastic and 15 kPa, n = 70 cells for 1.5 kPa; OCT4: n = 50 cells for plastic, n = 40 cells for 15 kPa and 1.5 kPa. k, Immunofluorescence assay for ETV4 in MCF-7 cells on PDMS layers with different stiffnesses. n = 45 cells. l, Immunofluorescence assay for ETV4 in hESCs after replating. n = 50 cells. n is number of cells (d,f,g,h,i,j,k,l) or regions (f) pooled from three independent experiments. Two-sided Student’s t-test, ***P < 0.001, **P < 0.01, *P < 0.05; ns, not significant. Exact P values are presented in Supplementary Table 9. Scale bars: 25 µm (c,g,h,i,j,k,l), 50 µm (f), 100 µm (a). Numerical source data and unprocessed gels are available as Source data.
To infer transcriptional regulators, we applied a recently developed analytic tool called Lisa (Landscape In Silico Deletion Analysis) that uses chromatic profile data26. Lisa analysis with top 200 downregulated DEGs in crowded hESCs (ordered by fold change) revealed 40 transcriptional regulators with significant P values (P < 0.01) and high expression in hESCs (fragments per kilobase of transcript per million mapped reads (FPKM) > 10; Fig. 1b and Supplementary Table 2). TEAD2 and SRF, previously known to be related to mechanotransduction, are included in the list. Protein function annotation with the selected transcription factors (TFs) using InterPro revealed seven protein domains with significant P values (adjP < 0.05). Among those identified, we found two terms related to PEA3-type ETS-domain TFs (Fig. 1b).
The PEA3 family consists of ETV1 (also known as ER81), ETV4 (also known as PEA3) and ETV5 (also known as ERM)27. Immunostaining validated the high expression of all PEA3 family TFs in hESCs and cell-crowding-induced reduction in protein expression (Fig. 1c–e). Among the PEA3 family TFs, we focused on ETV4 for its highest expression level in hESCs (Extended Data Fig. 1f). To test if cell-crowding-induced ETV4 downregulation is a reversible phenotype, we used a scratch assay, where rapid reactivation of ETV4 expression was observed in the cells adjacent to the scratches along with increased cell area (Fig. 1f). The cell-density-mediated regulation of ETV4 expression was also confirmed in other epithelial cell lines (Fig. 1g and Extended Data Fig. 1g). Direct manipulation of cell area by a cell stretching system revealed that a 15% decrease in cell area was sufficient to diminish the nuclear expression of ETV4 (Fig. 1h,i and Extended Data Fig. 1h–j).
Substrate stiffness is another physiologically relevant stimulus that regulates cellular mechanical stress28. Reduced mechanical stress by soft substrates was confirmed by immunostaining for pMLC and YAP/TAZ (Extended Data Fig. 1k,l). Compared with plastic (~1 Gpa), the nuclear expression of ETV4 proteins was significantly reduced when cells were placed on soft substrates (Fig. 1j,k). Replating hESCs to plastic resumed with high ETV4 expression (Fig. 1l). Overall, we have identified ETV4 as a TF whose expression is regulated by various mechanical cues.
Regulation of ETV4 expression by cell crowding dynamics
Like the in vivo epiblast, hESCs intrinsically grow, forming a single-layered epithelial colony in a culture dish (Fig. 2a). As hESC colonies expanded, we observed dynamic spatiotemporal changes in cell density. In small colonies (<0.5 mm2), individual cells exhibited a relatively large cell area, indicating a lesser degree of crowding (Fig. 2b). Conversely, in larger colonies (>2 mm2), cells located in the centre were smaller, while those at the periphery retained a larger cell area (Fig. 2b and Extended Data Fig. 2a). A gradual decrease in cell area was observed from the periphery to the centre of large hESC colonies (Fig. 2c). Because cell area exhibited a strong correlation with nucleus size (Extended Data Fig. 2b), we conducted real-time measurements of single nucleus sizes in an hESC line expressing a nuclear reporter, H2B-GFP. As the hESC colony expanded, an increase in single-cell variation in nucleus size was observed, with cells in the centre becoming smaller (Fig. 2d). These results collectively underscore the dynamic regulation of local cell crowding during epithelial expansion.
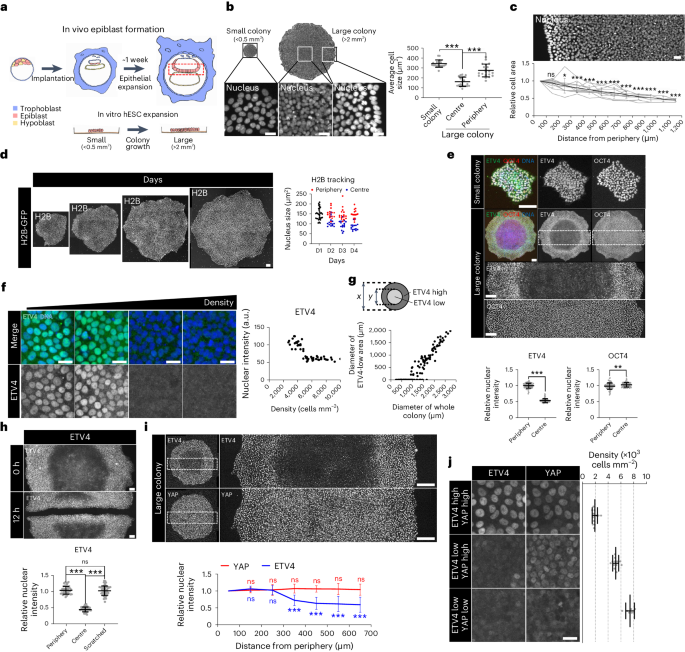
a, Schematic representation of in vivo epiblast formation and in vitro hESC expansion. b,c, Average cell area measured by dividing the total surface area by the number of cells. n = 20 regions (b) and n = 12 colonies (c). d, Time-course tracking assay for H2B-GFP in H9 colonies. n = 32 cells from three independent colony-tracking assays for H2B-GFP. e, Immunofluorescence assay for ETV4 and OCT4 in small and large H9 colonies. n = 80 cells. f, Immunofluorescence assay for ETV4 in different densities of hESCs. n = 52 cells. g, Quantitative analysis of the relationship between the diameters of ETV4-low areas and whole colonies in H9 hESCs. n = 168 colonies. h, Immunofluorescence assay for ETV4 in scratched H9 colonies. n = 50 cells. i, Immunofluorescence assay for ETV4 and YAP in large H9 colonies. Nuclear intensities for ETV4 and YAP signals were measured in single cells. n = 7 colonies. j, Cell density measurements in ETV4high/YAPhigh, ETV4low/YAPhigh and ETV4low/YAPlow regions of hESC colonies. n = 12 regions for ETV4high/YAPhigh and ETV4low/YAPhigh; n = 7 regions for ETV4low/YAPlow. n is number of cells (d,e,f,h), colonies (c,g,i) or regions (b) pooled from three independent experiments; or number of regions (j) pooled from four independent experiments. Two-sided Student’s t-test, ***P < 0.001, **P < 0.01, *P < 0.05; ns, not significant. Exact P values are presented in Supplementary Table 9. Scale bars: 25 µm (b,c,d,f,j), 50 µm (h), 100 µm (e) and 200 µm (i). Numerical source data are available as Source data.
Given the mechanosensitive regulation of ETV4 expression, we investigated the ETV4 expression patterns in H9 and H1 hESC colonies. ETV4 was homogeneously expressed in the nucleus of cells from small hESC colonies (<0.5 mm2; Fig. 2e and Extended Data Fig. 2c). Strikingly, large hESC colonies (>2 mm2) showed zonation of ETV4 expression with a reduction in the crowded centre (Fig. 2e and Extended Data Fig. 2c). However, the expression of core pluripotency genes (OCT4, NANOG and SOX2) remained high across the whole colony (Fig. 2e and Extended Data Fig. 2c,d). These results were confirmed in hESCs cultured either in a different medium or on a different coating extracellular matrix (ECM; Extended Data Fig. 2e,f). Clonal colonies derived from single hESCs also showed the zonation of ETV4 expression (Extended Data Fig. 2g). Consistent with the zonation pattern, sharp reduction in ETV4 expression was observed with increased cell density (Fig. 2f). The zonation pattern of ETV4 predominantly emerged when the colony diameter exceeded 1,000 μm, and the size of the ETV4-low area increased proportionally to the whole colony size (Fig. 2g). Reversible ETV4 activation in the centre of large colonies was confirmed by a scratch assay (Fig. 2h).
Because cell density influences various cellular processes, we first measured the cell shape index (CSI) to evaluate cell circularity at the boundary where ETV4 expression sharply transitions. High ETV4-expressing cells exhibited significantly larger cell areas than low ETV4-expressing cells, while both types of cells had similar CSIs (Extended Data Fig. 3a). These results suggest that ETV4 expression is associated with cell area rather than cell geometry. Next, we studied the potential effect of cell density on cellular metabolism. GSEA, based on the RNA-seq data from crowded hESCs, revealed no significant difference in the expression of glycolysis- and hypoxia-related genes (Extended Data Fig. 3b). To directly measure hypoxia, we used the hypoxyprobe system, in which pimonidazole hydrochloride forms protein adducts in hypoxic cells (Extended Data Fig. 3c)29. Consistent with the GSEA results, there was no evident induction of hypoxia in the crowded centre of large hESC colonies (Extended Data Fig. 3c). Furthermore, glucose uptake measurements demonstrated similar uptake rates between the periphery and the centre of large hESC colonies (Extended Data Fig. 3d,e). Although we cannot completely rule out the potential effect of cellular metabolism, these data indicate that cell-density-mediated ETV4 expression primarily depends on cell area.
Interestingly, crowded cells in the centre of large hESC colonies maintained active nuclear expression of YAP proteins despite the sharp inactivation of ETV4 (Fig. 2i and Extended Data Fig. 3f,g). We employed two shRNAs to target YAP (Extended Data Fig. 3h) and subsequently validated the accuracy of the YAP immunostaining results (Extended Data Fig. 3i). To gain deeper insights, we seeded hESCs at varying densities. Although nuclear expression of both ETV4 and YAP proteins was seen in a low density (~2,000 cells mm–2), inactivation of ETV4, but not YAP, took place in a medium density (~5,000 cells mm–2; Fig. 2j). At a high density (~7,500 cells mm–2), nuclear expression of both ETV4 and YAP proteins was reduced (Fig. 2j). The density ranges we used in this study are equivalent to those from other studies based on 2D gastruloid models6. These findings confirm that ETV4 and YAP respond differently to changes in cell density in hESCs. Given the crucial role of YAP/TAZ in epithelial proliferation10, a uniformly high YAP/TAZ level (Fig. 2i and Extended Data Fig. 3f,g) conforms to active cell division and high levels of the proliferation marker Ki67 and 5-ethynyl-2′-deoxyuridine (EdU) incorporation in the crowded centre (Extended Data Fig. 3j–l). YAP inhibition significantly diminished hESC colony growth (Extended Data Fig. 3m). Overall, these results demonstrate spatiotemporal regulation of ETV4 expression by cell crowding dynamics in a growing hESC epithelium.
Derepression of the neuroectoderm fate by ETV4 inactivation
Pluripotent epiblasts undergo dynamic changes in differentiation potential as the epithelial disc undergoes size expansion before gastrulation2,30. Consistently, growing evidence shows that the size of in vitro hESC colonies influences the differentiation propensity7,31,32, suggesting a direct role of epithelial expansion in lineage specification. Indeed, hESCs showed a biased differentiation toward mesendoderm (ME) when they were differentiated in a small colony. Under a robust neuroectoderm (NE)-directed differentiation condition called dual SMAD inhibition33, small colonies of hESCs were unable to produce NE cells expressing PAX6, which is a necessary and sufficient NE marker gene in humans (Fig. 3a and Extended Data Fig. 4a)34. However, ME-directed differentiation by BMP4 and FGF2 efficiently turned small hESC colonies into Brachyury+ ME cells (Fig. 3a and Extended Data Fig. 4a)35,36. In stark contrast, large hESC colonies produced both NE and ME lineage cells with clear spatial separation. PAX6+ NE cells predominantly emerged in the centre under dual SMAD inhibition, whereas ME cells were derived in the periphery region in the presence of BMP4 and FGF2 (Fig. 3a and Extended Data Fig. 4a). These results follow previous findings based on micropatterning technology6,7. The NE differentiation potential of large colonies was confirmed when hESCs were spontaneously differentiated by FGF2 and TGF-β deprivation (Extended Data Fig. 4b).
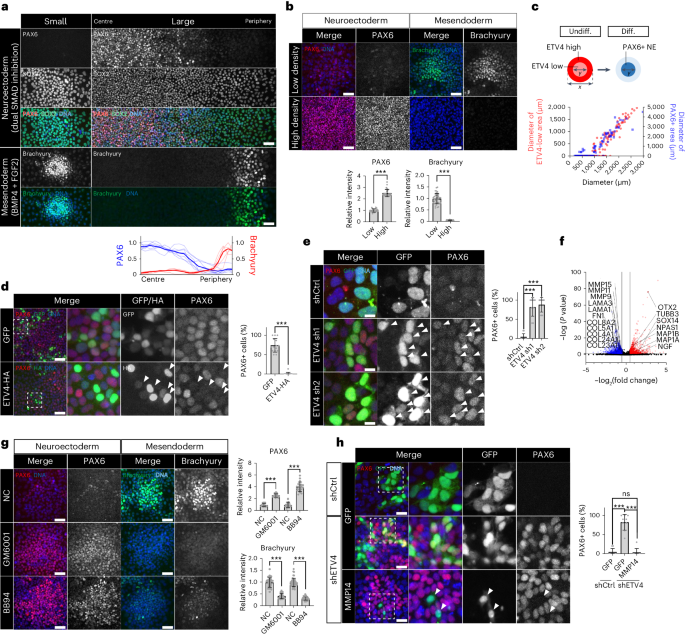
a, Immunofluorescence assay for PAX6, SOX2, Brachyury in differentiated H9 hESC colonies. n = 7 colonies. b, Immunofluorescence assay for PAX6 and Brachyury in H9 hESCs cells differentiated to either NE or ME cells. n = 25 regions for NE, n = 40 regions for ME. c, Quantitative analysis of the relationship between the diameters of ETV4-low areas in undifferentiated H9 hESC colonies and the diameters of PAX6+ areas in NE-differentiated colonies. n = 168 colonies for ETV4 and n = 92 colonies for PAX6. d, Immunofluorescence assay for PAX6 in large H9 colonies transduced with lentiviral vectors expressing ETV4-HA and differentiated to NE cells for 5 days. n = 15 regions. e, Immunofluorescence assay for PAX6 in small H9 colonies transduced with lentiviral vectors expressing ETV4 shRNAs together with GFP and differentiated to NE cells for 5 days. n = 10 regions. f, Volcano plot showing DEGs in H9 hESCs after ETV4 KD. The red and blue dots indicate upregulated and downregulated genes, respectively, with cutoff values for DEGs: log2(fold change) < −0.5 or > 0.5, adjP < 0.05. Full list of DEGs can be found in Supplementary Table 3. g, Immunofluorescence assay for PAX6 and Brachyury in small H9 colonies differentiated to either NE or ME cells with pan MMP inhibitors, GM6001 (5 μM) and BB94 (2 μM). NE differentiation: n = 21 regions for GM6001, n = 20 regions for BB94; ME differentiation: n = 38 regions for GM6001, n = 52 regions for BB94. h, Immunofluorescence assay for PAX6 in shETV4-expressing H9 hESCs transduced with lentiviral vectors expressing MMP14 together with GFP and differentiated to NE cells for 5 days. n = 9 regions. n is number of colonies (a,c) or regions (b,d,e,g,h) pooled from three independent experiments. Two-sided Student’s t-test, ***P < 0.001, **P < 0.01, *P < 0.05; ns, not significant. Exact P values are presented in Supplementary Table 9. Scale bars: 10 µm (e), 25 µm (h), 50 µm (a,d,g), 100 µm (b). Numerical source data are available in Source data.
Based on the above results, we hypothesized that the NE fate, initially repressed in small colonies, can be derepressed in the centre as cell crowding occurs during colony expansion. Indeed, high cell density dramatically promoted NE differentiation at the expense of ME derivation (Fig. 3b), consistent with the findings of a previous report33. Because ETV4 expression is downregulated in the crowded centre of large colonies, ETV4 could play a key role in suppressing the NE fate. The pre-patterns of ETV4 inactivation in undifferentiated hESC colonies precisely matched the size of the PAX6+ NE area in differentiated colonies (Fig. 3c). ETV4 expression was decreased in NE cells but not in ME cells, supporting the suppressive role of ETV4 in NE derivation (Extended Data Fig. 4c). Accordingly, ETV4 overexpression completely blocked the emergence of PAX6+ NE cells in large colonies (Fig. 3d and Extended Data Fig. 4d). Furthermore, ETV4 knockdown (KD) enabled small colonies to differentiate into NE cells (Fig. 3e and Extended Data Fig. 4e,f), which was blocked by ectopic ETV4 expression (Extended Data Fig. 4g). By contrast, ETV4 depletion impeded Brachyury+ ME cell differentiation (Extended Data Fig. 4h). Finally, cell crowding-mediated NE promotion was blocked by ETV4 overexpression (Extended Data Fig. 4i). These results demonstrate that ETV4 downregulation by cell crowding underlies NE lineage derepression.
To investigate the transcriptome-wide effect of ETV4 in hESCs, we performed RNA-seq after ETV4 KD (Fig. 3f and Supplementary Table 3). GO analysis of total DEGs (|log2(fold change)| > 0.5, adjP < 0.05) showed significant enrichment in embryonic development and morphogenesis in top 10 GO terms (Extended Data Fig. 5a), supporting the key role of ETV4 in early lineage determination. Moreover, DEGs upregulated by ETV4 KD included genes related to neural differentiation with nervous system development in TOP10 GO terms (Fig. 3f and Extended Data Fig. 5b), confirming the repressive role of ETV4 in the NE derivation. To pinpoint the molecular mechanisms of ETV4, we focused on downregulated DEGs because ETV4 primarily acts as a transcriptional activator37,38. Downregulated DEGs included many genes related to ECM remodelling, such as matrix metalloproteinases (MMPs; Fig. 3f), with ECM organization in top 10 GO terms (Extended Data Fig. 5c). Recently, it was reported that N-cadherin marks cells in the periphery of hESC colonies39. A re-analysis of published single-cell RNA-seq (scRNA-seq) data from sorted peripheral N-cadherin+ cells confirmed the elevated expression of a well-established ETV4 target gene, DUSP6 (Extended Data Fig. 5d,e)40. Furthermore, genes related to ECM remodelling, such as MMPs, were significantly upregulated in N-cadherin+ peripheral cells with ECM in top 10 GO terms (Extended Data Fig. 5e,f and Supplementary Table 4). These results support the role of ETV4 in ECM remodelling in the periphery region of hESC colonies.
ETV4 is a known direct upstream regulator of MMPs in cancer37,41,42,43. ETV4 KD or overexpression altered the expression of MMPs in hESCs (Extended Data Fig. 5g–i). The treatment of pan MMP inhibitors (GM6001 and BB94) was sufficient to derepress the NE fate in small hESC colonies (Fig. 3g). By contrast, MMP inhibition disrupted ME differentiation (Fig. 3g). Interestingly, membrane-type MMPs (MT-MMPs) such as MMP14 showed higher expression in hESCs than other MMPs (Extended Data Fig. 5j). The overexpression of MMP14 phenocopied ETV4 overexpression (Extended Data Fig. 5k,l) and blocked NE derepression by ETV4 KD (Fig. 3h). Overall, these results suggest that ETV4 is an NE repressor linking cell crowding dynamics to lineage specification.
Spatiotemporal ETV4 expression regulated by ERK
The MAPK signalling pathway is a well-known regulator of PEA3 family TFs in cancer44. To investigate this molecular link, we took advantage of the kinase translocation reporter (KTR) system45. When the kinase of interest is active, fluorescently-tagged substrates are phosphorylated and localized in the cytoplasm (Fig. 4a). The KTRs for ERK, p38 and JNK were validated by specific inhibitors (Extended Data Fig. 6a). For p38, individual cells displayed highly variable kinase activities with no discernible difference between the centre and periphery of large colonies (Extended Data Fig. 6b), whereas JNK activities were low in most cells (Extended Data Fig. 6c). However, the ERK-KTR showed clear cytoplasmic localization in most cells of small colonies (Fig. 4a). As a colony grew, a sharp reduction in ERK activity was observed in the crowded centre (Fig. 4b and Extended Data Fig. 6d,e). Induction of cell crowding was sufficient to inactivate ERK (Extended Data Fig. 6f). Live cell imaging of ERK activity revealed that the ERK activity pattern closely resembled the expression pattern of ETV4 (Fig. 4c).
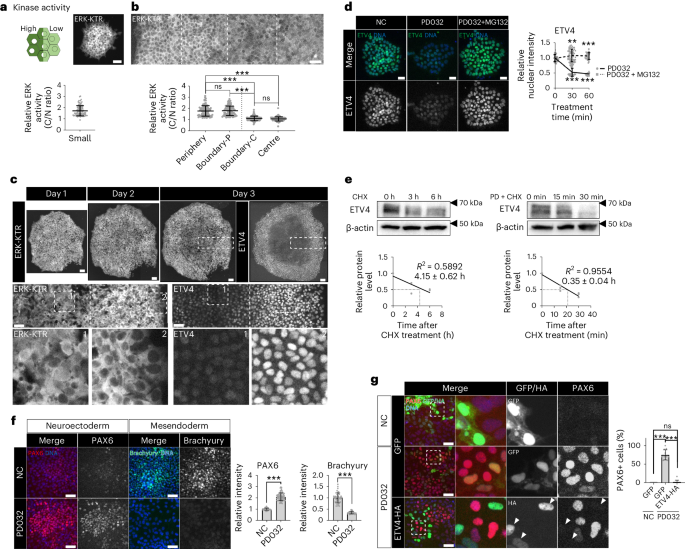
a,b, ERK activity measured by the ratio of the cytoplasmic over nuclear intensities of ERK-KTR (C/N ratio) in single cells within small (a) and large (b) H9 colonies. n = 95 cells for small; n = 85 cells for periphery and centre; n = 90 cells for boundary-P (peripheral side near the boundary where ERK acivity transitions) and boundary-C (central side near the boundary). c, Representative time-course images from three independent colony-tracking assays for ERK-KTR in H9 colonies, followed by immunofluorescence assay for ETV4. d, Immunofluorescence assay for ETV4 in H9 hESCs treated with PD0325901 (1 μM) and MG132 (10 μM). n = 60 cells. e, Quantification of ETV4 protein stability in H9 hESCs in the presence of PD0325901 (1 μM). For CHX, n = 3 (0 h), 3 (3 h) and 2 (6 h) independent experiments. For PD + CHX, n = 3 independent experiments. PD, PD0325901; CHX, cycloheximide. f, Immunofluorescence assay for PAX6 and Brachyury in small H9 colonies differentiated to either NE or ME cells with PD0325901 (1 μM). NE differentiation: n = 75 regions for NC, n = 69 regions for PD0325901; ME differentiation: n = 42 regions. g, Immunofluorescence assay for PAX6 in H9 hESCs transduced with lentiviral vectors expressing ETV4-HA and differentiated to NE cells for 5 days with PD0325901 (1 μM). n = 11 regions. n is number of cells (a,b,d) or regions (f,g) pooled from three independent experiments. Two-sided Student’s t-test, ***P < 0.001, **P < 0.01, *P < 0.05; ns, not significant. Exact P values are presented in Supplementary Table 9. Scale bars: 25 µm (d), 50 µm (a,b,f,g), 100 µm (c). Numerical source data, unprocessed gels and additional microscope images are available as Source data.
ERK inhibition by chemical inhibitors (PD0325901 and U0126) induced a rapid decrease in ETV4 protein abundance without affecting the mRNA level (Fig. 4d and Extended Data Fig. 6g,h). Proteasome inhibition blocked the decrease, suggesting that ERK regulates ETV4 protein stability (Fig. 4d). ERK inhibition dramatically reduced the half-life of ETV4 proteins from 4.15 h to 0.35 h (Fig. 4e). COP1 is a critical E3 ligase for the degradation of PEA3 family TFs46,47. In large hESC colonies, COP1 was primarily localized in the nucleus with homogenous expression (Extended Data Fig. 7a). COP1 KD slightly increased the basal level of ETV4 protein (Extended Data Fig. 7b–d) and nullified the effect of ERK inhibition on ETV4 expression (Extended Data Fig. 7d). COP1 depletion also blocked ETV4 downregulation in the crowded centre of large hESC colonies and suppressed NE differentiation (Extended Data Fig. 7e,f).
Finally, ERK inhibition derepressed the NE fate and inhibited ME differentiation in small hESC colonies (Fig. 4f). NE derepression by ERK inhibition was completely blocked by ETV4 overexpression (Fig. 4g). ERK activation by constitutively active KRASG12V increased ETV4 protein levels and suppressed NE differentiation (Extended Data Fig. 7g–i). While ERK signalling is known to have a broad impact on numerous regulatory factors, our findings suggest ETV4 as a primary target of ERK in the context of lineage specification.
Cell-crowding-mediated regulation of receptor endocytosis
The FGF and TGF-β signalling pathways play crucial roles in maintaining pluripotency48,49,50,51. Short-term treatment of A83-01 (TGF-β inhibitor) showed no effect on ERK activity (Fig. 5a); however, SU-5402 (FGF inhibitor) reduced ERK activity and the level of ETV4 proteins without altering mRNA expression (Fig. 5b,c). We used shRNAs targeting FGFR1 owing to the high expression in hESCs (Extended Data Fig. 8a–c). FGFR1 KD decreased ERK activity and ETV4 expression (Extended Data Fig. 8d,e), suggesting FGFR1-mediated signalling as an upstream pathway of ERK and ETV4.
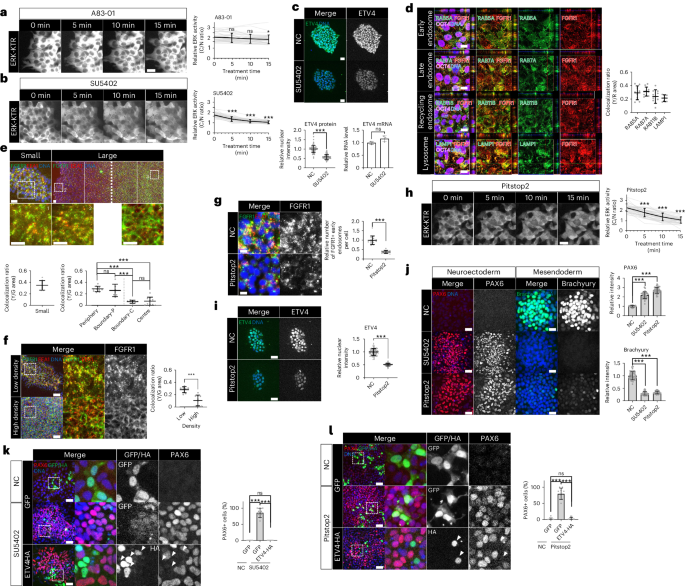
a,b, Time-course images for ERK-KTR in H9 hESCs treated with A83-01 (5 μM; a) or SU5402 (10 μM; b). ERK activity was measured by the ratio of the cytoplasmic over nuclear intensities. n = 35 cells. c, Immunofluorescence assay and qPCR analysis for ETV4 in H9 hESCs treated with SU5402 (10 μM, 1 h). n = 77 cells for immunofluorescence assay and n = 3 samples for qPCR. d, Immunofluorescence assay for FGFR1 in H9 hESCs expressing various reporters. Colocalization was measured by the yellow/red (Y/R) ratio. n = 9 regions for RAB5A and LAMP1, n = 10 regions for RAB7A and RAB11B. e, Immunofluorescence assay for FGFR1 and EEA1 in small and large H9 colonies. Colocalization was measured by the yellow/green (Y/G) ratio. n = 8 regions for small, n = 10 regions for periphery and centre, n = 7 regions for boundary-P and boundary-C. f, Immunofluorescence assay for FGFR1 and EEA1 in H9 hESCs in low- and high-density cultures (24 h after seeding). Colocalization was measured by the Y/G ratio. n = 9 regions for low density and n = 10 regions for high density. g, Immunofluorescence assay for FGFR1 and EEA1 in H9 hESCs treated with Pitstop2 (50 μM, 30 min). n = 15 regions for NC and n = 13 regions for Pitstop2. h, Time-course images for ERK-KTR in H9 hESCs treated with Pitstop2 (50 μM). n = 40 cells. i, Immunofluorescence assay for ETV4 in H9 hESCs treated with Pitstop2 (50 μM, 30 min). n = 120 cells. j, Immunofluorescence assays for PAX6 and Brachyury in small H9 colonies differentiated to either NE or ME cells with SU5402 (10 μM) or Pitstop2 (20 μM). n = 40 regions. k, Immunofluorescence assay for PAX6 in small H9 colonies transduced with lentiviral vectors expressing ETV4-HA and differentiated to NE cells for 5 days with SU5402 (10 μM). n = 9 regions. l, Immunofluorescence assay for PAX6 in small H9 colonies transduced with lentiviral vectors expressing ETV4-HA and differentiated to NE cells for 5 days with Pitstop2 (20 μM). n = 9 regions. n is number of cells (a,b,c,h,i) or regions (d,e,f,g,j,k,l) pooled from three independent experiments. Two-sided Student’s t-test, ***P < 0.001, **P < 0.01, *P < 0.05; ns, not significant. Exact P values are presented in Supplementary Table 9. Scale bars: 10 µm (d,g), 25 µm (a,b,c,e,h,i,j), 50 µm (f,k,l). Numerical source data are available as Source data.
An emerging body of evidence shows that receptor localization contributes to regulating downstream signalling52,53. For example, it was recently reported that lateral localization of TGF-β receptors impeded cellular responses to apically applied ligands6,54. However, we observed that cell-crowding-induced ETV4 inactivation occurred in hESC culture on a transwell system, where ligands are accessible to both apical and basolateral sides of cells (Extended Data Fig. 8f). In the case of receptor tyrosine kinases, endocytosis is functionally related to the downstream signalling activation52. Activated EGFR and FGFR accumulate in endosomes that serve as a signalling platform55, and blocking receptor endocytosis inhibits the downstream signalling56,57,58. Recently, it was reported that membrane tension influenced FGFR endocytosis and downstream ERK activity in mouse ESCs, implicating mechanical regulation of receptor endocytosis59,60. To test this idea, we generated stable hESC lines expressing GFP reporters for various endosomes and lysosomes61. In small hESC colonies, a significant number of FGFR1-containing vesicles colocalized with RAB5A+ early endosomes (Fig. 5d), a finding that was confirmed using another early endosome marker, EEA1 (Fig. 5e). FGFR1-containing vesicles were also observed in late endosomes (RAB7A), recycling endosomes (RAB11B) and lysosomes (LAMP1; Fig. 5d). These results suggest that FGFR1 proteins are under active endocytosis, followed by recycling or degradation. Strikingly, cells in large hESC colonies showed distinct subcellular localizations of FGFR1 proteins depending on their positions. These proteins were predominantly present in endosome vesicles in the periphery of large colonies (Fig. 5e and Extended Data Fig. 9a). In stark contrast, membrane localization of FGFR1 proteins was observed in the crowded centre, suggesting impaired endocytosis (Fig. 5e and Extended Data Fig. 9a, b). FGFR1 proteins were localized in both apical and basolateral membranes in the crowded centre (Extended Data Fig. 9a). The transition of FGFR1 localization occurred abruptly along the radial axis of large hESC colonies (Fig. 5e). To further validate these results, we applied a DNA aptamer (TD0) previously developed as an agonist of FGFR1 (Extended Data Fig. 9c)62. Fluorophore-tagged aptamers were found in FGFR1-containing early endosomes in the periphery of large hESC colonies (Extended Data Fig. 9d). By contrast, few aptamer-containing early endosomes were detected in the centre (Extended Data Fig. 9d). Furthermore, induction of cell crowding was sufficient to impair FGFR1 endocytosis with loss of EEA1 colocalization (Fig. 5f). These results show that FGFR1 endocytosis is tightly regulated by cell crowding.
Next, we used Pitstop2 (a clathrin inhibitor) and Dynasore (a dynamin inhibitor) to suppress endocytosis (Extended Data Fig. 9e)63,64. Endocytosis inhibition dramatically decreased the number of FGFR1-containing endosomes, ERK activity and ETV4 expression (Fig. 5g–i and Extended Data Fig. 9f–h), which phenocopied the effects of FGFR inhibition (Fig. 5b,c). Constitutively active KRASG12V blocked the effect of endocytosis inhibition on ETV4 expression (Extended Data Fig. 9i), suggesting that endocytosis regulates ETV4 through the FGFR–RAS signalling.
Consistent with previous research65,66, we found that FGFR inhibition was sufficient to derepress the NE fate within small hESC colonies (Fig. 5j and Extended Data Fig. 9j), while suppressing the ME derivation (Fig. 5j). Notably, we observed a similar phenotypic outcome upon the inhibition of endocytosis (Fig. 5j). Furthermore, ectopic ETV4 expression entirely counteracted the NE-promoting effects induced by FGFR and endocytosis inhibitors (Fig. 5k,l). Overall, our findings suggest that cell-crowding-mediated blockade of FGFR1 endocytosis leads to the inactivation of both ERK and ETV4.
Regulation of receptor endocytosis by integrin–actomyosin
To figure out molecular mechanisms bridging cell crowding to FGFR1 endocytosis, we revisited the RNA-seq data performed in crowded hESCs. Downregulated DEGs in crowded hESCs showed significant enrichment of genes related to cell–ECM interactions such as integrins (Fig. 6a and Extended Data Figs. 1b and 10a). Published scRNA-seq data from N-cadherin+ peripheral cells also showed focal adhesion in top 10 KEGG pathways (Extended Data Fig. 10b)39. These results concord with the fact that cell crowding leads to a smaller surface area through which cells interact with the ECM.
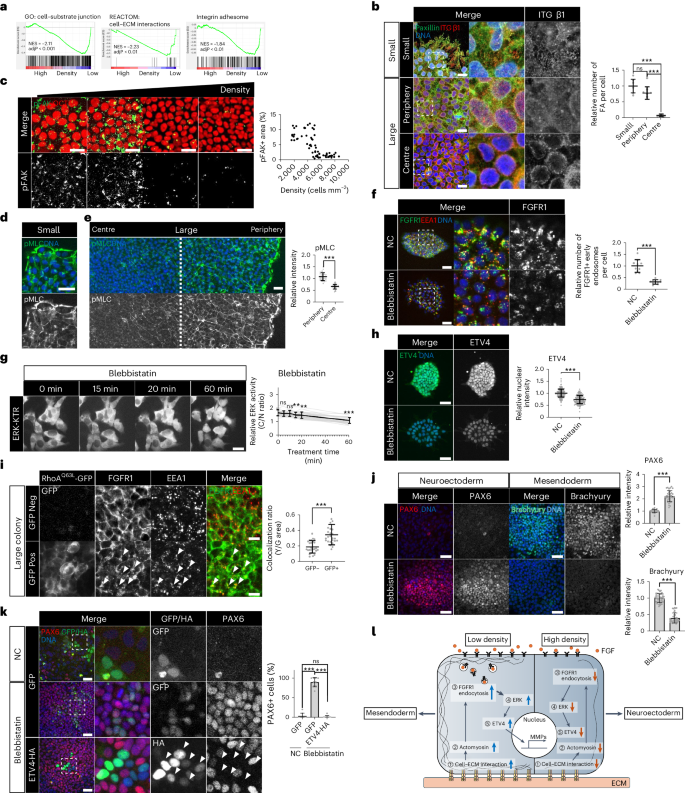
a, GSEA with the RNA-seq data from H9 hESCs in low- and high-density cultures. GSEA was performed with three different gene sets: cell–substrate junction (left), cell–ECM interaction (middle) and integrin adhesome (right). b, Immunofluorescence assay for Integrin β1 and Paxillin in small and large H9 colonies. n = 9 regions for small, n = 13 regions for periphery, and n = 10 regions for centre. c, Immunofluorescence assay for pFAK and OCT4 in different densities of H9 hESCs. n = 69 regions. d,e, Immunofluorescence assay for pMLC in small (d) and large (e) H9 colonies. n = 30 regions. f, Immunofluorescence assay for FGFR1 and EEA1 in small H9 colonies treated with Blebbistatin (50 μM, 30 min). n = 10 regions. g, Time-course images for ERK-KTR in H9 hESCs treated with Blebbistatin (50 μM). ERK activity was measured by the ratio of the cytoplasmic over nuclear intensities (C/N ratio). n = 30 cells. h, Immunofluorescence assay for ETV4 in H9 hESCs treated with Blebbistatin (50 μM, 1 h). n = 241 cells for NC and n = 230 cells for Blebbistatin. i, Immunofluorescence assay for FGFR1 and EEA1 in large H9 colonies transduced with lentiviral vectors expressing constitutively active RhoAQ63L-GFP in a doxycycline-dependent manner. n = 25 regions. j, Immunofluorescence assay for PAX6 and Brachyury in small H9 colonies differentiated to either NE or ME cells with Blebbistatin (1 μM). NE differentiation: n = 37 regions, ME differentiation: n = 45 regions for NC and n = 38 regions for Blebbistatin. k, Immunofluorescence assay for PAX6 in H9 hESCs transduced with lentiviral vectors expressing ETV4-HA and differentiated to NE cells for 5 days with Blebbistatin (1 μM). n = 9 regions. l, Graphic summary of the mechanotransduction pathway regulating ETV4 expression and lineage fates. n is number of cells (g,h) or regions (b,c,e,f,i,j,k) pooled from three independent experiments. Two-sided Student’s t-test, ***P < 0.001, **P < 0.01, *P < 0.05; ns, not significant. Exact P values are presented in Supplementary Table 9. Scale bars: 10 µm (b,i), 25 µm (c,d,e,f,g), 50 µm (h,j,k). Numerical source data are available as Source data.
Consistent with the transcriptomic results, a high level of integrin β1 was observed in paxillin+ focal adhesions in small colonies and the periphery of large colonies (Fig. 6b). By contrast, crowded cells in the centre showed perinuclear localization of integrin β1 with loss of paxillin colocalization (Fig. 6b). Focal adhesion kinase (FAK) is a critical component of integrin signalling67. Immunostaining of phosphorylated FAK (pFAK, active FAK) revealed the sharp downregulation in response to increased cell density in hESCs (Fig. 6c). Furthermore, FAK inhibition suppressed FGFR1 endocytosis, ERK activity and ETV4 expression (Extended Data Fig. 10c–f). These findings suggest that integrin signalling bridges cell crowding to FGFR1 endocytosis.
Although integrin signalling has the potential to impact a wide array of cellular processes, we focused on its role in regulating cytoplasmic actin filaments, given the significance of actin dynamics in receptor endocytosis68,69,70. Consistent with the patterns of integrin β1+ focal adhesions, small hESC colonies showed well-established pMLC+ bundles, while the zonation of pMLC staining appeared in large hESC colonies with a sharp reduction in the centre (Fig. 6d,e). Disassembly of pMLC+ bundles was observed by integrin inhibition (RGDS peptide), FAK inhibition or cell crowding (Extended Data Figs. 1e and 10g,h). We used either blebbistatin (myosin inhibitor) or YM (Y-27632, RhoA kinase inhibitor; ML-7, myosin light chain kinase inhibitor) to inhibit actomyosin (Extended Data Fig. 10i,j)71. Transient inhibition of actomyosin activity decreased the number of FGFR1-containing endosome vesicles and inactivated ERK and ETV4 (Fig. 6f–h and Extended Data Fig. 10d,k,l). Importantly, the transient activation of actomyosin by doxycycline-inducible expression of constitutively active RhoAQ63L partially re-activated FGFR1 endocytosis in the crowded centre of large hESC colonies (Fig. 6i and Extended Data Fig. 10m). These findings suggest that actomyosin, in addition to serving as a marker of cellular mechanical stress, plays a crucial role in regulating receptor endocytosis in response to mechanical stimuli. However, it is essential to acknowledge that other cellular changes induced by cell crowding, such as the reduction in apical surface area for ligand interaction and alterations in cell geometry, may also contribute to regulating receptor endocytosis.
Finally, actomyosin inhibition derepressed the NE fate in small hESC colonies while preventing ME differentiation (Fig. 6j). The NE derepression by actomyosin inhibition was blocked by ETV4 overexpression (Fig. 6k). Overall, these results suggest that the integrin–actomyosin pathway serves as a crucial link connecting cell crowding to FGFR1 endocytosis, ETV4 expression and NE specification (Fig. 6l).
Dynamic ETV4 expression captured by mathematical modelling
A critical question that remains to be answered in this study is how the gradient of cell crowding from the periphery to the centre produces a sharp boundary in ETV4 expression. To address this question, we propose a mathematical framework that describes a cell crowding model. Based on real-time imaging data of H2B-GFP hESC colony growth, we obtained an analytical solution to the governing reaction-diffusion partial differential equation describing cell crowding dynamics (Methods). This equation assumes a cell population density varying in time and space due to the flow of cells (diffusion) from high- to low-density regions (outward flow) as well as cell division (reaction). A similar approach has been used to describe bacterial colony growth72. Our model accurately predicted the colony population and colony size (diameter; Fig. 7a). Moreover, the model predicted an increase in cell density over time and higher cell crowding at the colony centre (Fig. 7b).
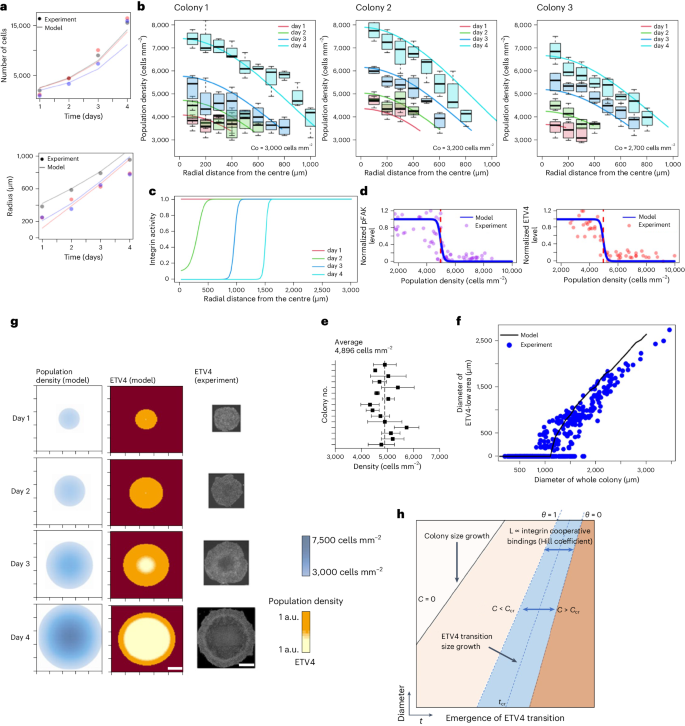
a, Model (solid lines) for hESC colony growth that accurately predicts colony growth and proliferation dynamic. Different colours represent three different colonies. b, Comparison between model and observed spatiotemporal change in cell population density. Solid lines show model prediction and boxplots represent the range of experimentally measured cell population densities at certain distance on days 1, 2, 3 and 4. The three panels illustrate three different colonies with an initial average cell population density C0. The black lines in the middle of the boxes are the median values for each group. The vertical size of the boxes illustrates the interquartile range. Whiskers represent 1.5 × interquartile range. n = 4 regions within a colony for each distance. c,d, Model prediction for integrin activity (c), pFAK and ETV4 (d) transition that occurs at a critical cell population density of ~5,000 cells mm–2. Simulation generated for an example colony with an initial radius of 250 μm and an initial average cell population density 3,000 cells mm–2. e, Critical cell densities for ETV4 inactivation measured across H9 colonies. n = 3 regions within a colony. f, Comparison between model and experimental measurement for ETV4 transition (Fig. 2g). Simulation generated for an example colony with an initial radius of 250 μm and an initial average cell population density 3,000 cells mm–2. n = 168 colonies pooled from three independent experiments. g, Model for spatiotemporal change of population density and radial distribution of ETV4. Scale bar, 600 μm. Simulation generated for an example colony with an initial radius of 250 μm and an initial average cell population density 3,000 cells mm–2. h, ETV4 transition map as a function of space and time.
Cell crowding inversely correlates with ECM-accessible area, linking ECM ligand availability to cell density. Moreover, it is well established that integrin–ECM binding kinetics follow a cooperative interaction73,74. Such an interaction enforces a system with an on/off switch-like behaviour. We used the Hill equation to model integrin–ECM interactions75. The equation captures the biomolecular interaction that exhibits the cooperativity between two binding molecules. Our model predicts a switch for integrin activity that occurs at different distances from the colony’s centre as the colony grows over time (Fig. 7c). The ultrasensitive transition of integrin activity appears when the core cell density reaches a critical density of 5,000 cells mm–2 (Fig. 7d), which matches well with the pFAK and ETV4 experimental data (Figs. 2f, 6c and 7e). Furthermore, our model recapitulated the experimental data with the ETV4 low-expression region appearing in the centre when the colony size reaches 1,000–1,500 μm in diameter (Fig. 7f,g). The critical cell density can only be observed after t > tcr (where t represents time and tcr represents the critical time when the ETV4 transition boundary appears for the first time), and the radius at which the ETV4 transition occurs varies as a function of time (Fig. 7h). Although our model successfully demonstrated the ultrasensitive transition of integrin activity and ETV4 expression based on the ECM availability and the cooperativity in integrin–ECM interaction, it is essential to acknowledge that cell shape itself may also contribute to regulating integrin signalling or the downstream pathways independently of ECM availability76. Overall, our model bridges the simple growth of stem cell epithelium to the timing and location of ETV4 inactivation and NE derepression.
- SEO Powered Content & PR Distribution. Get Amplified Today.
- PlatoData.Network Vertical Generative Ai. Empower Yourself. Access Here.
- PlatoAiStream. Web3 Intelligence. Knowledge Amplified. Access Here.
- PlatoESG. Carbon, CleanTech, Energy, Environment, Solar, Waste Management. Access Here.
- PlatoHealth. Biotech and Clinical Trials Intelligence. Access Here.
- Source: https://www.nature.com/articles/s41556-024-01415-w