High-purity isolation of mature OP9 adipocytes
The bone marrow (Fig. 1A) is a complex tissue with diverse cellular species. In addition to hematopoietic cells, it includes MKs and BMAds, both characterized by their large size and fragility in suspension. To develop our sorting device, we first utilized OP9 cells, a non-clonal line of bone marrow-derived mouse stromal cells known as a robust adipogenesis model47,48. After in vitro adipocytic induced differentiation, OP9 progenitors accumulated lipid droplets and underwent the expected size increase (Fig. 1B, in suspension). Figure 1C shows pre- and post-differentiation OP9 cultures. The differentiation outcome depended on passage number and confluency at induction, thus induced-OP9 samples resulted in varying ratios of differentiation stages, from progenitors to mature adipocytes, ranging from 7 μm to 40 μm in size.
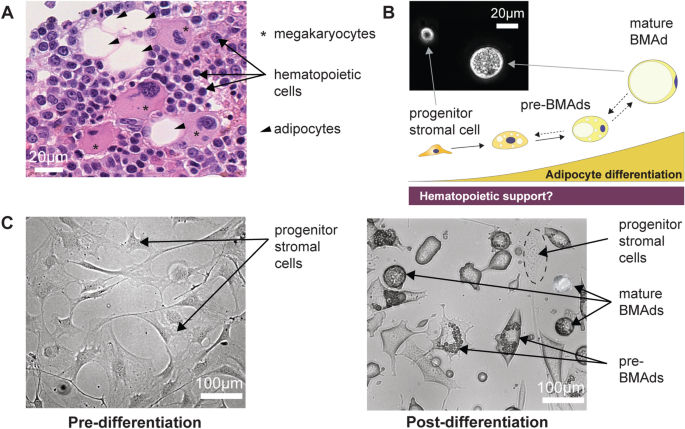
Bone Marrow niche and Heterogeneous Adipocytic Differentiation in vitro. (A) Hematoxylin and eosin-stained slide of human bone marrow (BM) trephine biopsy imaged at 40X magnification using a Nanozoomer S60, revealing hematopoietic cells (labeled with arrows), megakaryocytes (labeled with asterisks), and bone marrow adipocytes (BMAds) (labeled with arrow heads). (B) Schematic representation of the BMAd differentiation axis as a study model for the relationship between hematopoiesis and adipogenesis. Inset: phase contrast imaging of cells in suspension at the progenitor and mature BMAd stages, highlighting the size difference between these two populations. (C) OP9 progenitor cells seeded at 20,000 cells/cm2 (undifferentiated-OP9) show a homogeneous fibroblastic-like cell structure in adherent cultures. Following in vitro induced adipocytic differentiation (induced-OP9, 6 days post-differentiation), the sample contains various stages of maturation: progenitors (fibroblast morphology), pre-BMAds (cells with limited lipid droplet accumulation), and mature BMAds (round cells filled with lipids).
The microchip layout and SEM images of the sorting module are presented in Fig. 2A and 2B, and supplementary videos (SV1–2) show the microchip operation. Figure 2C and 2D depict the behavior of microbeads within MarrowCellDLD and the sorting workflow for adipocytic cultures. Figure 3A shows a sample of OP9-derived adipocytic culture (induced-OP948), transiting the MarrowCellDLD sorter at the inlet and outlet. Specifically, a single-cell suspension was continuously injected and focused using lateral sheath flows (Fig. 3A, inlet). Stirring the sample prevented bias due to cell buoyancy and an embedded filtering system at the inlet tube reduced doublets and clusters (Fig. 2D). Cluster removal was essential to guarantee cell purity and continuous sample processing for up to 3–4 h without clogging issues.
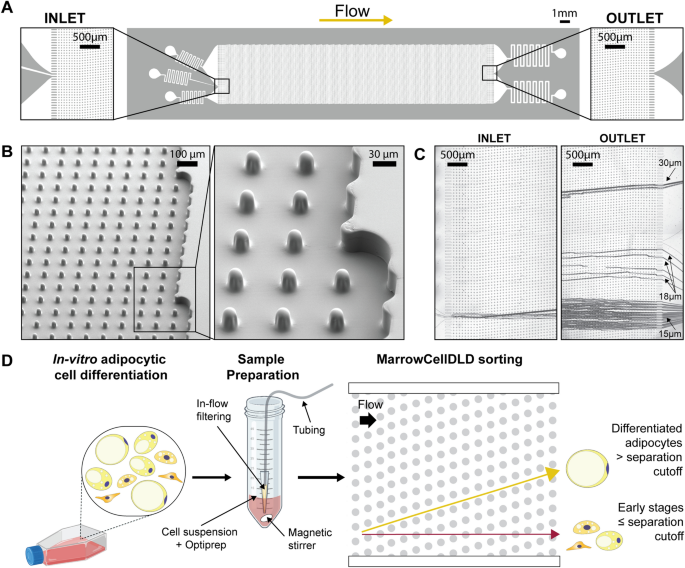
MarrowCellDLD device and operation. (A) MarrowCellDLD chip: insights on inlet and outlet regions terminated with 10 rows of straight pillars respectively before and after the MarrowCellDLD array active region (tilted array). (B) Scanning electron microscopy images of the MarrowCellDLD array chip with a 19 μm separation cutoff (critical size). (C) Inlet and outlet trajectories of polystyrene microbeads of 15, 18, and 20 μm in size transiting a MarrowCellDLD chip with 19 μm critical size. (D) Experimental workflow to sort differentiated adipocytes by MarrowCellDLD. After adipocytic differentiation in vitro, the induced-OP9 sample contains a mixture of progenitor cells, early stages of differentiation, and mature adipocytes. After trypsinization, the cellular sample is suspended in its original culture media supplemented with Optiprep, then placed at the inlet reservoir connected to the tubing responsible for injecting the sample into the MarrowCellDLD device. A custom in-flow filtering system embedded in the tubing ensures the injection of a single-cell suspension, and the sample is continuously stirred to achieve homogeneity. Within the sorting module, mature adipocytes larger than the critical size for separation should be forced to follow the array angle (displacement mode), allowing for their isolation. Conversely, early stages of differentiation and progenitors should move parallel to the flow (zig–zag mode). The two fractions are thus predicted to be physically separated and can be collected at distinct outlets.
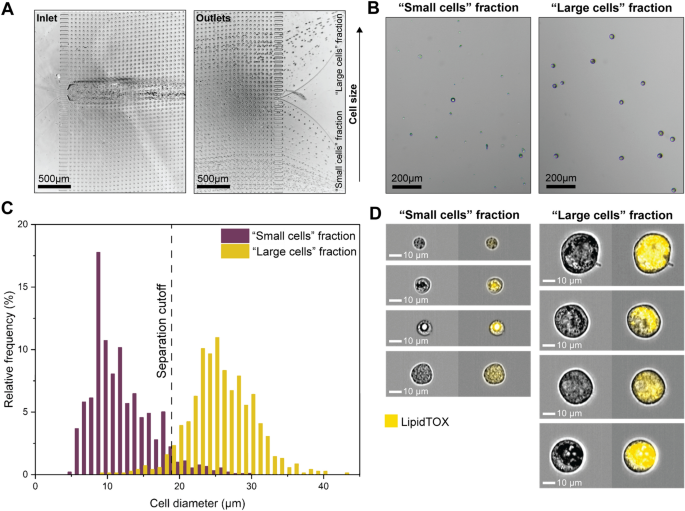
Fluid Dynamic MarrowCellDLD Sorting of Induced-OP9 Cells. (A) Trajectories of cells entering (inlet) and exiting (outlets) the microfluidic system. The test involved injecting induced-OP9 cells after 6 days of differentiation at a concentration of 500,000 cells/mL. (B) Phase contrast microscopy of outlet fractions after MarrowCellDLD sorting an induced-OP9 mixture. (C) Cell diameter distributions of outlet fractions (« small cells » fraction, violet; « large cells » fraction, yellow) after sorting induced-OP9s by MarrowCellDLD with a 19 µm separation cutoff. Phase contrast microscopy of cells collected at the outlets and replated after sorting as in (B) enabled cell size quantification in QuPath0.3.2. Data are displayed as relative frequencies over 150 cells per fraction from n = 4 sorting experiments. (D) ImageStream flow cytometry imaging (brightfield and LipidTOX-stained) of cells from the two fractions separated by MarrowCellDLD. MarrowCellDLD sortings were performed with a 19 µm separation cutoff and applied pressure of 20 mbar.
The MarrowCellDLD sorting array was designed to retrieve two fractions based on a separation cutoff size determined by the array periodicity and gap between pillars. The injected induced-OP9 single-cell suspension comprised a continuous distribution of cell sizes. As expected from the design, cells exited the array spreading across the width of the main channel, which subdivides downstream into two subchannels denominated « small cells » and « large cells » fractions (Fig. 3A, outlets). Notably, the « small cells » outlet collected more events per unit time, including small debris and contaminations, whereas the « large cells » outlet was highly purified (Fig. 3B).
We tested MarrowCellDLD sorting arrays with different separation cutoffs (DLD critical sizes). The nominal separation cutoff was calculated based on the general DLD model38. We initially designed devices at nominal critical sizes 15, 17.5, 20, 22.5, and 25 μm and fabricated them as PDMS replicas. The gaps between the pillars in the fabricated DLD arrays measured by surface profilometry were larger than the designed ones, therefore the actual separation cutoffs resulted larger than the nominal critical sizes. We first characterized the sorting modules by processing polystyrene microbeads of different sizes (Fig. 2C). Subsequently, we tested the five different array geometries with induced-OP9 samples and observed the sorted fractions by phase contrast microscopy. Nominal critical sizes of 22.5 μm and 25 μm were too large for retrieving mature OP9 adipocytes. We found that a nominal critical size of 15 μm corresponded to a 19 μm separation cutoff, which was ideal for isolating mature OP9 BMAds (Fig. 3C). The gap of 42 ± 1 μm in this array was sufficient to preserve intact mature adipocytes with minimal blockage (few units per sorting run).
Specifically, Fig. 3C shows the size distributions of the two output fractions after sorting induced-OP9s by MarrowCellDLD with a 19 µm separation cutoff. Cells collected at the outlet reservoirs were imaged by phase contrast microscopy, and QuPath0.3.2 software49 was used for cell diameter quantification (n = 4 independent experiments with 150 cells per fraction). We found that 97 ± 2% of cells within the « large cells » fraction were above the 19 μm cutoff, representing an approximately three-fold enrichment compared to the original induced-OP9 adipocytic sample (with only 38% above 19 μm). Consistently, the « small cells » fraction contained 96% ± 3% of cells below the separation cutoff. Independent sorting experiments through different MarrowCellDLD devices showed reproducible purity for both fractions, independent from the original mixture composition. The combined size distribution of the sorted fractions matches that of the original sample, indicating the preservation of cell size dynamics. Additionally, we further characterized devices with a nominal critical size of 20 μm, as they could provide the finest separation among the ones tested in our study. Figure S1 shows the sorting outcome for a device with a nominal critical size of 20 μm that corresponded to an actual separation cutoff of 24 μm, achieving 90–93% purity over the « large cells » fraction for two independent sorting experiments. It is worth noting that with both geometries we could reliably retrieve intact BMAds above 35 μm in diameter, the size of the largest adipocytes found in the murine BM, as defined within the intact tissue.
To validate our findings, we observed the populations sorted by MarrowCellDLD by ImageStream flow cytometry imaging (Fig. 3D). LipidTOX, a lipophilic stain commonly used for adipocyte FACS sorting, was used to tag the lipid-laden cells and LipidTOX signals quantified by Imagestream analysis of the sorted fractions (Fig. S2). Indeed, all cells within the « large cells » fraction exhibited positive LipidTOX staining (98% ± 2%, n = 3), indicating significant lipid accumulation typical of mature BMAds. Large adipocytes and diverse levels of lipid drop coalescence were observed. Interestingly, several cells in the « small cells » fraction (63% ± 12%, n = 3) also displayed LipidTOX positivity at varying intensities. Some pre-BMAds showed small lipid droplets coalescing, producing a significant LipidTOX signal even if not fully mature. These results suggest that BMAds sorting based solely on lipid staining intensity may not be sufficient to differentiate between adipocyte maturation stages. Gently purifying adipocyte populations with a precisely defined size range can help to overcome this limitation.
Phenotype of sorted induced-OP9 adipocyte fractions
In addition to ImageStream flow cytometry imaging, BODIPY staining and phase contrast microscopy were used to interrogate the phenotype and neutral lipid content of the cells in the small and large cell fractions as compared to the unsorted mix (Fig. S3A). To confirm the presence of mature BMAds in the « large cells » fraction and morphologically discern BMAds from their progenitors or intermediately differentiated cells, we employed a BODIPY fluorescence stain, indicative of cellular lipidic content and thus of the extent of adipocyte maturation upon differentiation. Compared to the unsorted mixture comprising 81% BODIPY-positive cells, we found the great majority (95%) of cells in the « large cells » sorted fraction to stain positive for BODIPY (Fig. S3C-D). Conversely, a small fraction (18%) of cells within the « small cells » fraction was stained for BODIPY, confirming the enrichment of smaller, non-lipidated cells (Fig. S3B). Overall, we concluded that MarrowCellDLD sorting of induced-OP9 adipocytic cell suspensions could successfully separate large, lipidated, intact BMAds at high purity from unlipidated or adipogenesis-refractory precursors as determined by the phenotype and lipid content of the sorted cells.
Viability and functionality of sorted induced-OP9 adipocytes
Next, we evaluated the viability of induced-OP9s before and after MarrowCellDLD sorting (n = 15, with approx. 100 cells per experiment) (Fig. 4A). Cell viability was measured using Trypan Blue and a hemocytometer. Sample filtering and Optiprep addition did not significantly influence cell viability (pre-sorting viability: 91% ± 7%; post-sample preparation viability: 91% ± 8%). Cells remaining in the input vial after the sorting process exhibited comparable viability (unsorted leftover viability: 86% ± 8%). As for the sorted cells, a significant but modest decrease in viability (82% ± 6%) was observed as compared to the pre-sorting sample, but the difference was no longer significant when compared to the unsorted leftover. Overall, viability never fell below 70% for all sorting experiments performed.
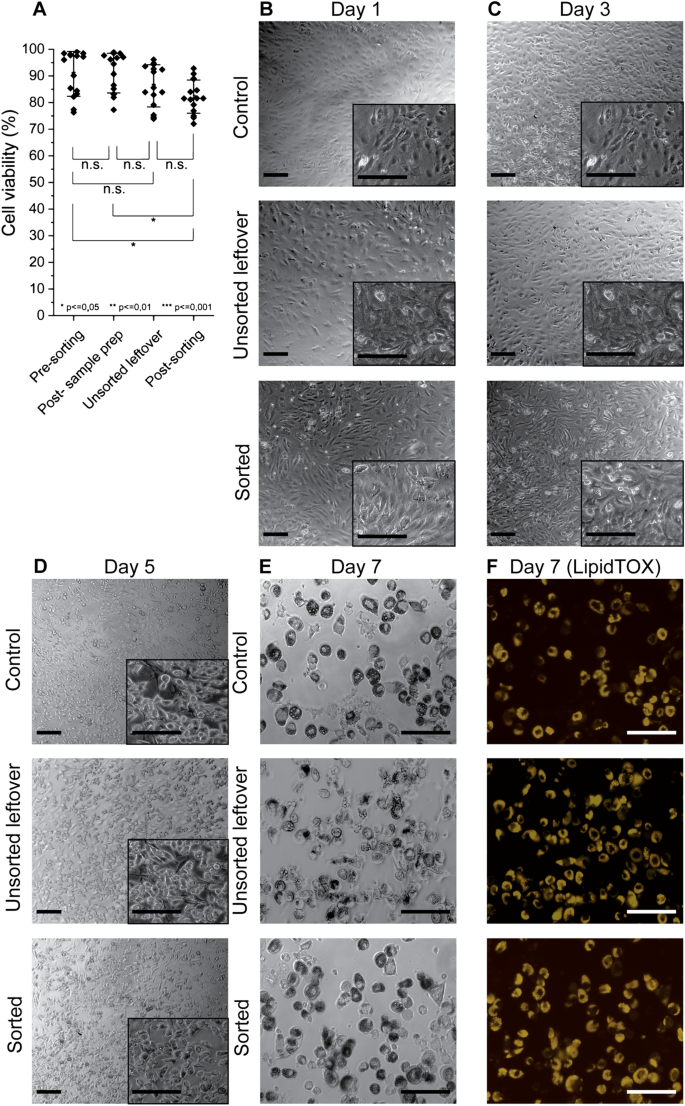
Cell viability and functionality of OP9 cells after MarrowCellDLD sorting. (A) Cell Viability measurements: (i) pre-sorting, (ii) post-sample preparation, (iii) unsorted leftover, and (iv) cells collected at outlets post-sorting (small and large cell fractions). Statistical significance was evaluated by the Student’s t-test for independent samples (n = 15 MarrowCellDLD sorting experiments on induced-OP9 samples). (B–F) Time-sequential images at different days post-plating following MarrowCellDLD sorting of undifferentiated OP9 cells. Phase contrast and fluorescence (Yellow, LipidTOX staining) images for control cells (unsorted), unsorted leftover (unsorted sample residual at the inlet reservoir after 3 h sorting), and sorted (collected at the «small cells » fraction outlet) after induced adipocytic differentiation for 6 days. Scale bar: 100 µm. MarrowCellDLD sortings were performed with a 19 µm separation cutoff and applied pressure of 20 mbar.
As shown in Fig. 4B–F, we further assessed post-sorting viability and functionality by comparing (i) undifferentiated OP9 cells that were subject to neither sample preparation nor sorting (Control), (ii) undifferentiated-OP9 cells remaining unsorted at the input vial (Unsorted leftover), and (iii) sorted undifferentiated OP9 progenitor cells (Sorted). Undifferentiated cultures were chosen for these experiments to ensure adherence for downstream differentiation assays, as otherwise, the low adherence displayed by mature adipocytes from induced-OP9 cultures would have made the sorted cell fractions very difficult to compare. Time-lapse phase contrast microscopy and fluorescence (LipidTOX) images were captured on days 1, 3, 5, and 7. Notably, the sorted OP9 cells, unsorted leftover cells, and control cell fractions all adhered and proliferated at days 1 and 3, which further confirms post-sorting viability. To determine functionality, we induced differentiation of the three samples. Subsequent imaging on days 5 and 7 revealed clear evidence of adipocytic differentiation, confirmed by LipidTOX staining, which reflected lipid droplet accumulation. Therefore, neither the sorting nor the sample preparation and stirring affected the ability of OP9 progenitors to differentiate in BMAds. Overall, we could thus validate the reliable performance of MarrowCellDLD in isolating different phenotypes of induced-OP9 adipocytic cells with preserved viability and functionality post-sorting.
Comparison with FACS sorting
We compared the performance of MarrowCellDLD with FACS sorting, the gold-standard approach for precise cell sorting at high throughput. FACS sorting relied on LipidTOX intensity to isolate adipocytic cultures into three distinct populations after gating on single viable cells (PI-negative or DAPI-negative), classified respectively as Low, Medium, and High LipidTOX fractions. Note that the FACS-sorting gates for the High LipidTOX fractions were inclusive of all detectable FSC/SSC high events. This gating approach (Fig. S4A-B) was implemented employing two different FACS instruments: BD FACSAria™ III (BD Biosciences) at a nozzle pressure of 20 psi and MoFlo Astrios EQ (Beckman Coulter) at a reduced pressure of 10 psi. Lower pressure is expected to preserve fragile adipocytes but implies a slower sorting process.
Following FACS sorting, all output fractions were analyzed by ImageStream to extract the diameter of single viable cells (Fig. S4C). Figure 5 shows the sorting outcomes with Aria and Astrios FACS instruments, with cell size distributions in Fig. 5A and 5C, respectively. Representative ImageStream micrographs of the MarrowCellDLD fractions are shown in Fig. 5B and 5D, as compared to Fig. 3C. A single FACS sorting experiment is directly compared with a single MarrowCellDLD sorting of induced-OP9s with the same passage number, to compare samples with similar size dynamics. The unsorted cells were subjected to the same sample preparation protocols as the sorted fractions and kept at the input reservoirs for the whole process for both MarrowCellDLD and FACS sortings.
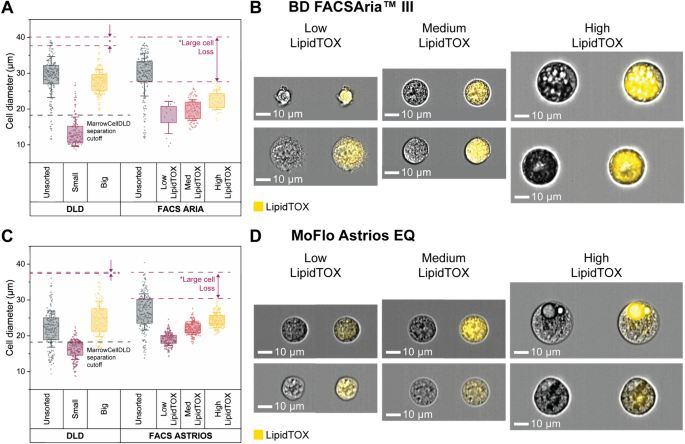
Comparison between MarrowCellDLD and FACS sorters BD FACSAria™ III and MoFlo Astrios EQ. MarrowCellDLD size-based separation is compared to FACS sorting based on LipidTOX staining for lipidic content. (A, C) Post MarrowCellDLD or FACS sorting, cells from the unsorted sample and the sorted fractions were imaged by ImageStream flow cytometry. Distributions of sizes of single viable cells of each fraction after sorting by MarrowCellDLD, FACS Aria, and MoFlow Astrios FACS instruments. Cell diameters are extracted by ImageStream analysis for each method (200 cells per group or total cells analyzed in the fraction plotted from a single experiment, selected from n = 2 biological replicates using BD FACSAria™ III cell sorter and n = 3 biological replicates using MoFlo Astrios EQ cell sorter). Data are displayed with standard deviation as error bar, and box plot representing the 25% to 75% data points of the total sample. (B, D) Representative ImageStream images of brightfield and LipidTOX-stained cells from the FACS sorted fractions obtained by FACS Aria and MoFlo Astrios, respectively. MarrowCellDLD sortings were performed with a 19 µm separation cutoff and applied pressure of 20 mbar.
For the FACS Aria versus MarrowCellDLD comparison, unsorted cells were highly differentiated, with 75% cells above 26 μm (Fig. 5A and 5B). MarrowCellDLD effectively isolated cells above the predetermined 19 μm cutoff, purifying mature adipocytes and even preserving cells larger than 35 μm. Conversely, the high LipidTOX fraction obtained after FACS Aria sorting surprisingly lacked cells above 30 μm. Notably, we did not observe any intact unilocular adipocytes after sorting by FACS Aria. Despite ImageStream confirming the higher degree of lipid accumulation within the high LipidTOX fraction, cells in the other two sorted populations were to a lesser extent positive for the lipophilic stain and overlapped in terms of size. We thus concluded that LipidTOX gating alone was insufficient to discriminate between different degrees of lipidation and that the vast majority of large mature adipocytes was lost during the FACS Aria sorting process.
We then moved to compare the MarrowCellDLD device to the low-sorting-pressure MoFlo Astrios FACS instrument. We included FSC/SSC gating into the FACS-sorting strategy to better discriminate the Low and Medium LipidTOX populations13,47,50. For this set of experiments, the unsorted induced-OP9 samples in the MarrowCellDLD experiment exhibited a narrower range of cell sizes (Fig. 5C), therefore including a lower proportion of large, mature adipocytes than for Fig. 5A. As shown in Fig. 5C, upon ImageStream analysis we found that MarrowCellDLD isolated cells above the 19 μm separation cutoff and preserved cells larger than 35 μm. For the MoFlow Astrios FACS-sorted fractions, we observed LipidTOX-positive cells in all three fractions, and a harmonious increase in size proportional to the lipid signal (Fig. 5D). The MoFlo Astrios sorter could retrieve more intact viable high-LipidTOX cells than the FACS Aria instrument, but again produced losses of large high-LipidTOX cells (Fig. 5C). Specifically, cells within the MoFlo Astrios high-LipidTOX fraction displayed lipid droplet accumulation but often did not exhibit complete differentiation, and unilocular adipocytes were not observed (Fig. 5D). We, therefore, concluded that contrary to the MarrowCellDLD device, and although less damaging, the low-pressure MoFlow Astrios FACS sorting still lacked the gentleness required to preserve all fragile adipocytes.
Then, we compared the MarrowCellDLD performance to the MoFlow Astrios FACS instrument in terms of processing time and yield (Table 1). While MarrowCellDLD is label-free, FACS requires LipidTOX staining incubation (30 min) and washing (15 min) steps before sorting. Additionally, centrifugation for medium exchange to a FACS buffer potentially harms the cells and introduces buoyancy-based biases. For a sample of 1.5 × 106 cells, i.e. the sample size of induced-OP9 cells in a T25 culture flask, MarrowCellDLD required 180 min of sorting time, while FACS Astrios approximately 120 min. Including in the total processing time the sample preparation times, which for FACS include staining incubation times, the FACS process required 170 min in total and MarrowCellDLD took approximately 185 min, both within the same order of magnitude.
To estimate losses of large adipocytes, we compared the size distributions of sorted and unsorted cells (Fig. 5C). MoFlow Astrios FACS sorting could not recover 50% of cells larger than 19 μm in the original sample, calculated as the percentage of unsorted cells above the mean + standard deviation in the high LipidTOX fraction. In contrast, for MarrowCellDLD, the comparison between the large cell fraction and the unsorted population revealed that only 7% of the large cells were lost during sorting. Finally, we should note that FACS does not entail a dilution of the original sample, while MarrowCellDLD, at this stage of our design, introduces a 40-fold dilution.
Finally, we tested the effectiveness of isolating large lipidated cells from the induced-OP9 single-cell suspensions using the floatation-based protocol developed to isolate primary adipocytes from femoral surgical debris, introduced by Attané et al.18. Unfortunately, and as reported by the authors (personal communication) we were not able to obtain a floating layer of high-buoyancy adipocytes from our in vitro differentiated murine stromal cultures even after long waiting times post-centrifugation (Supplement Fig. S5), as opposed to the primary human BMAds their technique was developed for. The expected cell diameter heterogeneity of induced-OP9 adipogenic cultures was retrieved within the pelleted fraction, including numerous cells larger than 35 μm. Therefore, although we could not compare the buoyancy method for mature adipocyte isolation, we could demonstrate that centrifugation enables the concentration of mature OP9-derived adipocytes to counteract the dilution introduced by MarrowCellDLD.
MarrowCellDLD sorting of spontaneous OP9, induced MSOD, and induced megakaryocytes
MarrowCellDLD consistently demonstrated high-performance sorting of induced-OP9 adipocytes. To further validate the versatility of our device for fragile cell types, we conducted MarrowCellDLD sorting experiments on two additional adipogenesis models: spontaneously differentiated OP9 cells, where mature adipocytes are rare (Fig. 6A) and adipogenesis-induced MSOD human-derived stromal cells (Fig. 6B), as well as human megakaryocytes derived from primary CD34+ cells (Fig. 6C). For the adipocyte models, we followed the same sample preparation protocol and sorted an equal number of cells (1.5 × 106 cells) in each scenario.
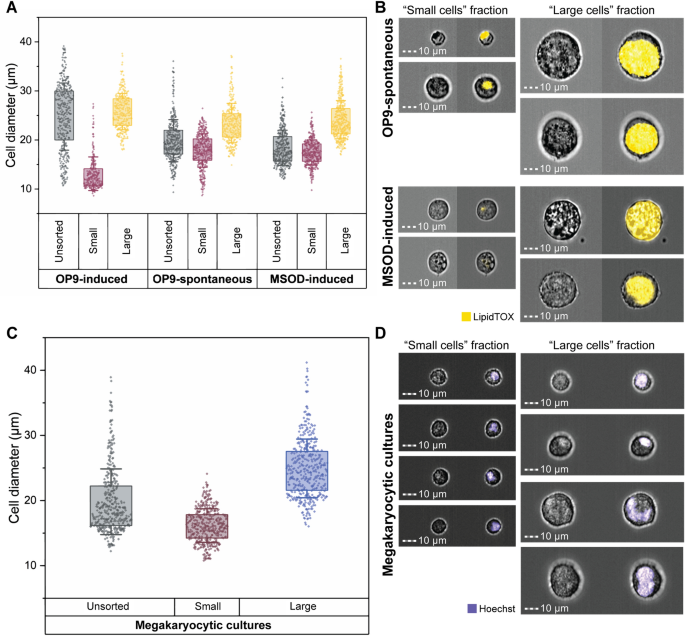
MarrowCellDLD sorting of different BM cell types. We compared the MarrowCellDLD sorting of adipocytes from (i) induced (OP9-induced) or (ii) spontaneously differentiated (OP9-spontaneous) mouse-derived OP9 stromal cells, as well as (iii) human-derived MSOD progenitors (MSOD-induced). (A) After MarrowCellDLD sorting, cells were stained by DAPI and imaged by ImageStream flow cytometry to quantify the diameter of DAPI-negative single cells (n = 2 experiments for each cell type, with 400 cells per group). (B) Representative ImageStream micrographs of OP9-spontaneous and MSOD-induced cells (brightfield and LipidTOX-stained) separated by MarrowCellDLD sorting onto « small cells » (left panels) and « large cells » (right panels) and outlet based on a predefined critical size cutoff of 19 μm. (C) MarrowCellDLD sorting of human megakaryocytes derived from primary CD34+ hematopoietic progenitor cells. Cells were stained by Hoechst and imaged by ImageStream flow cytometry, to quantify the diameter of the single Hoescht-positive cells (n = 2 experiments, with 400 cells per group). (D) Representative ImageStream micrographs upon MarrowCellDLD sorting of megakaryocytic cultures, showing megakaryocytes on the « large cells » fraction (brightfield and Hoechst-stained after MarrowCellDLD sorting). MarrowCellDLD sortings were performed with a 19 µm separation cutoff and applied pressure of 20 mbar.
As expected, the post-differentiation single cell-mixture of spontaneous-OP9s, or induced-MSODs, consistently included fewer mature BMAds as compared to the previously tested induced-OP9 samples. However, even if both OP9- and MSOD-derived mature BMAds were rare in these conditions, the MarrowCellDLD device successfully separated at high purity large cells above the 19 μm separation cutoff (Fig. 6A). Specifically, spontaneous-OP9 adipocytes were collected at 90% purity and induced-MSOD adipocytes at 94% purity. ImageStream assessment of the LipidTOX-labeled fractions confirmed that cells within the « small cells » fraction corresponded to the phenotypes of unlipidated progenitor cells or early-stage lipidated cells, while fully-lipidated cells with mature BMAd phenotype were collected within the « large cells » fraction (Fig. 6B). This correlation was evident from both the intensity and area of the LipidTOX stain, as well as the cell size inferred from the brightfield images. Thus, we can conclude that MarrowCellDLD could be also used for high-purity sorting of rare BMAds, in vitro-derived from both mouse and human progenitor cells. Notably, the largest adipocytes (> 35 μm) were preserved even if extremely rare.
Finally, we tested our method to isolate differentiated megakaryocytes from primary human progenitors. Differentiation of human CD34+ hematopoietic stem and progenitor cells isolated from peripheral blood was induced in vitro. Upon megakaryocytic differentiation, progenitors undergo endomitosis resulting in large, polyploid cells, expressing the characteristic surface markers CD41 and CD42b9. The extent of differentiation is largely donor-dependent. Similar to OP9 and MSOD cell lines, the post-differentiation sample contained a mixture of progenitors and various stages of differentiation. Large cells within this mixture, as defined by FSC on flow cytometry and by size assessment (diameter > 19 µm) by ImageStream, were double positive for the megakaryocyte-specific markers CD41 and CD42b (Supplement. Fig. S6). We tested the capacity of the MarrowCellDLD device (19 µm separation cutoff) to sort hematopoietic cell mixtures after megakaryocytic differentiation (Fig. 6C). Cells above the separation cutoff were isolated at 96% purity and retrieved intact, including cells larger than 35 µm in diameter. Congruently, we found higher polyploidy, as measured by DNA dye Hoechst 33,258, in the « large cells » fraction as compared to the « small cells » fraction of the respective MarrowCellDLD outlets (Fig. 6D). Cell viability was not significantly affected, neither by maintaining the sample under agitation during the process nor by the MarrowCellDLD sorting procedure itself (Supplement. Fig. S7). Overall, our results confirm the robustness and adaptability of MarrowCellDLD as a powerful sorting method for various fragile cell types, based on a predefined size cutoff, including terminally differentiated cells from human sources.
- SEO Powered Content & PR Distribution. Get Amplified Today.
- PlatoData.Network Vertical Generative Ai. Empower Yourself. Access Here.
- PlatoAiStream. Web3 Intelligence. Knowledge Amplified. Access Here.
- PlatoESG. Carbon, CleanTech, Energy, Environment, Solar, Waste Management. Access Here.
- PlatoHealth. Biotech and Clinical Trials Intelligence. Access Here.
- Source: https://www.nature.com/articles/s41598-023-47978-w